IASR 42(11), 2021【THE TOPIC OF THIS MONTH】Inf luenza 2020/21 season, Japan
PDF download(PDF:42.63KB)
The topic of This Month Vol.42 No.11(No. 501)![]()
Inf luenza 2020/21 season, Japan
(IASR Vol. 42 p239-241:November 2021)
In the 2020/21 season (week 36 in September 2020 to week 35 in August 2021), the number of reported inf luenza cases did not increase during the usual epidemic period of the calendar year. Multiple other indicators also remained at remarkably low levels, and no signs suggestive of an inf luenza epidemic were observed. This may have been due to the epidemic of coronavirus disease 2019 (COVID-19) and countermeasures against it.
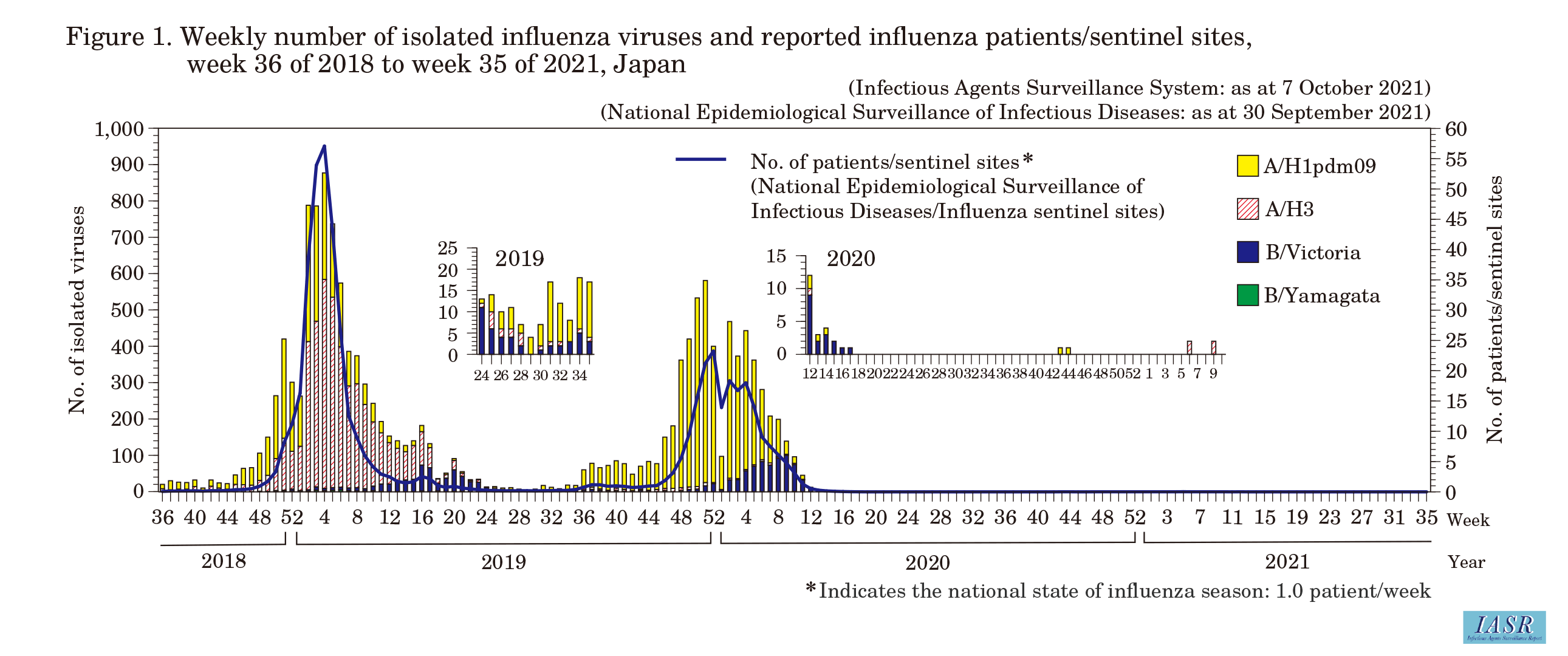
Epidemiology of the 2020/21 inf luenza season: Under the National Epidemiological Surveillance of Infectious Diseases (NESID) system, approximately 5,000 inf luenza sentinel sites nationwide (approximately 3,000 pediatric and 2,000 internal medicine sentinel sites) report patients diagnosed with inf luenza on a weekly basis. In the 2020/21 season, the maximum number of reported inf luenza cases per sentinel was 0.02/week (98 patients reported/week) in the f ifth week of 2021, and no week exceeded 1.00/week, which is the threshold for the nationwide start of the inf luenza season (Fig. 1) (see p. 242 of this issue). Based on the reported sentinel surveillance data, the estimated cumulative total number of medically-attended inf luenza patients nationally was approximately 14,000 (week 36 of 2020 to week 17 of 2021), which was substantially lower than that during the same period of the past two seasons (7,289,000 patients in the 2019/20 season and 12,005,000 patients in the 2018/19 season). The total number of hospitalized patients in the hospitalized inf luenza patient surveillance (data from approximately 500 designated sentinel hospitals with at least 300 beds) was also extremely low at 131 patients in the 2020/21 season (week 36 of 2020 to week 17 of 2021), compared with 20,719 patients in the 2018/19 season and 13,011 patients in the 2019/20 season. The number of schools with class closures reported to the “inf luenza-like illness surveillance” system (school closure and absenteeism surveillance) was 3 schools (week 36 of 2020 to week 9 of 2021), a drastic decrease from the same period of the last two seasons when more than 20,000 schools with class closures were reported. Among patients diagnosed with acute encephalitis (including encephalopathy), a category V notif iable infectious disease, there were no cases diagnosed as inf luenza encephalopathy. No signs of an inf luenza epidemic were observed through the inf luenza-like illness surveillance system conducted in Mie Prefecture (see p.243 of this issue).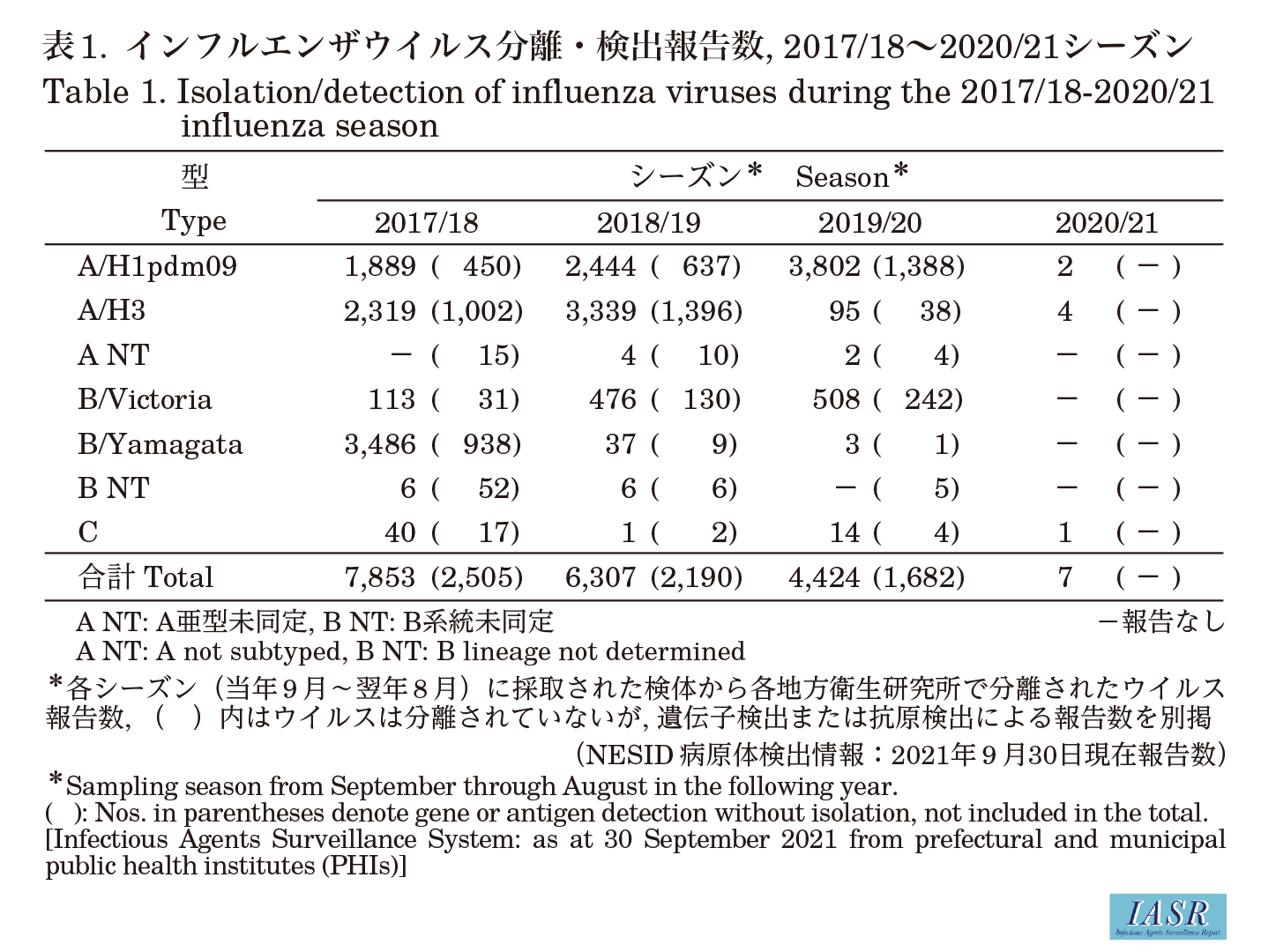

Isolation/detection of inf luenza virus in the 2020/21 season: Prefectural and municipal public health institutes reported the isolation/detection of 7 inf luenza viruses in the 2020/21 season (Table 1). Among them, 5 were reported from inf luenza sentinel sites and 2 from non-sentinel sites (Table 2). Regarding isolates/detections of inf luenza, two subtype A/H1pdm09 viruses and four subtype A/H3 viruses were reported, but there were no reports of type B virus for either the Victoria or the Yamagata lineage. One type C inf luenza virus was detected (Table 2). Two subtype A/H1pdm09 viruses were reported from Nagasaki Prefecture in weeks 43 and 44 of 2020, two subtype A/H3 viruses were reported from Yamagata Prefecture in week 6, and another two subtype A/H3 viruses were reported from Miyagi Prefecture in week 9 of 2021 (Fig. 1 and Fig. 2).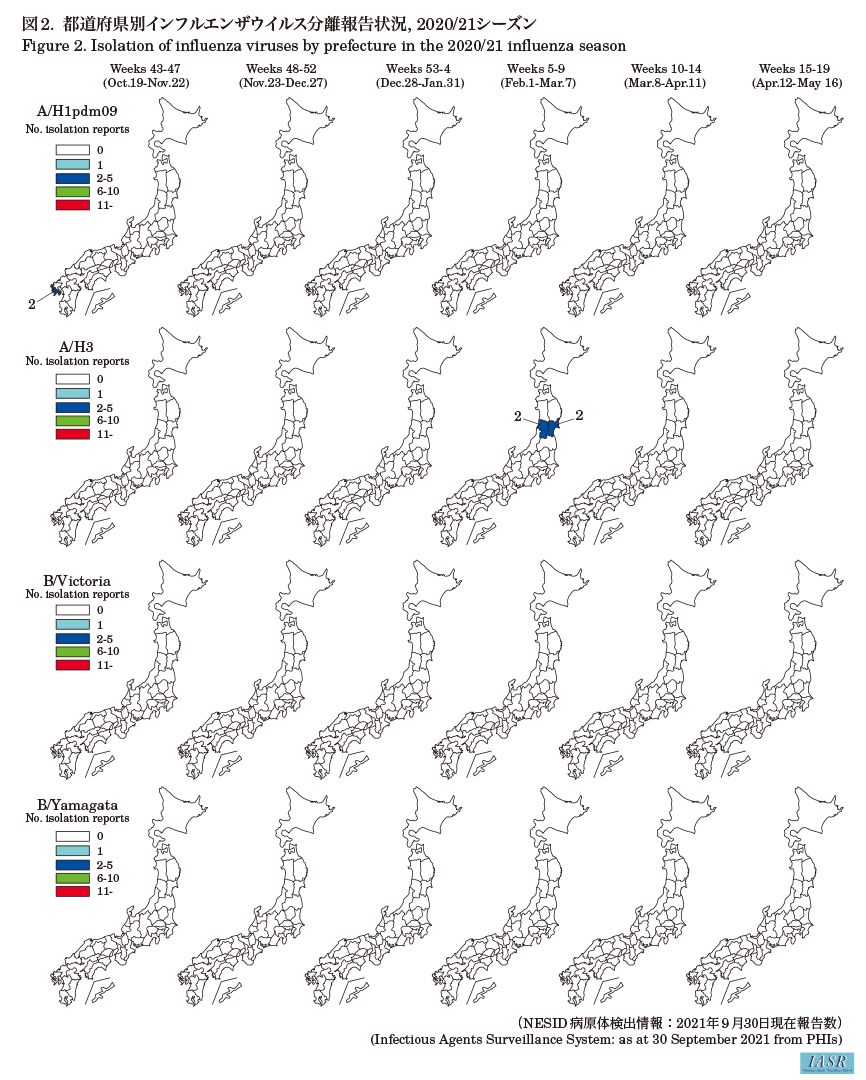
Genetic and antigenic characteristics of the 2020/21 isolates: The National Institute of Infectious Diseases (NIID) conducted genetic and antigenic analyses of isolates from Japan and other Asian countries. Antigenic analyses were done using ferret antiserum (see p. 247 of this issue).
The two subtype A/H1pdm09 strains detected in Japan were classif ied into genetic subclade 183P-5A1 (D187A, Q189E) based on the phylogenetic tree analysis of the hemagglutinin (HA) gene. Antigenic analysis revealed that the two isolates reacted well with ferret antiserum against the “egg-isolated” A/Guangdong-Maonan/SWL1536/2019 (183P-5A1), the vaccine strain for the 2020/21 season.
For subtype A/H3, phylogenetic analysis of the HA gene of four domestic strains and 14 isolates from Laos and Nepal showed that the domestic and Laotian strains belonged to subclade 3C.2a1b.2a1 and the Nepalese strain belonged to subclade 3C.2a1b.2a2. Antigenic analysis showed that all tested strains had poor reactivity with ferret antiserum against the “cell culture propagated” A/Hong Kong/45/2019 and “egg-isolated” A/Hong Kong/2671/2019, the vaccine strains for the 2020/21 season that belong to subclade 3C.2a1b.1b.
Although the type B/Victoria lineage virus was not detected in Japan, the HA gene phylogenetic tree analysis of the strains detected overseas showed that most of the strains belonged to subclade 1A.3a1 or 1A.3a2. In antigenic analysis, these viruses did not react well with ferret antiserum against “cell culture propagated” or “egg isolated” B/Washington/02/2019, the vaccine strain for the 2020/21 season. No viruses of the type B/Yamagata lineage were analyzed.
Resistance to antivirals among the 2020/21 isolates: The two subtype A/H1pdm09 viruses isolated in Japan during the 2020/21 season were not resistant to neuraminidase (NA) inhibitors (oseltamivir, peramivir, zanamivir, and laninamivir) or a cap-dependent endonuclease inhibitor (baloxavir), but were resistant to amantadine. The 14 subtype A/H3 viruses isolated in Japan and overseas (Nepal and Laos) were not resistant to NA inhibitors or a cap-dependent endonuclease inhibitor, but were resistant to amantadine. No strains of type B virus were available for testing (see p.247 of this issue).
Seroprevalence among the Japanese population prior to the 2020/21 season: Based on seroprevalence surveys conducted under the National Epidemiological Surveillance of Vaccine-Preventable Diseases program, seroprevalence levels were measured from 3,244 serum samples collected between July to September 2020 (see p.252 of this issue). The age groups with the highest seroprevalence (measured as HI antibody-positive, titer ≥1:40) were 10-24-year-olds (≥ 40%) for A/Guangdong-Maonan/SWL1536/ 2019 [subtype A/H1pdm09 vaccine strain], 5-49- and ≥ 70-year-olds (41-83%) for A/Hong Kong/2671/2019 [subtype A/H3 vaccine strain], 15-39-year-olds (≥ 60%) for B/Phuket/3073/2013 [B/Yamagata lineage vaccine strain], and 40-54-year-olds (30-40%) for B/Victoria/705/2018 [B/Victoria lineage vaccine strain]. Antibody prevalence against A/Hong Kong/2671/2019 [A/H3 subtype vaccine strain] was the highest among the four strains analyzed.
Inf luenza vaccine: The tetravalent vaccine for the 2020/21 season contained antigens of two type A and two type B strains. Approximately 33,420,000 vial-equivalent doses (estimated on the assumption that 1 vial contained 1 mL) were produced, of which an estimated 32,740,000 vials were used.
For the 2021/22 season, A/Victoria/1/2020 (IVR-217) was selected as the vaccine strain for subtype A/H1pdm09 and A/Tasmania/503/2020 (IVR-221) as the vaccine strain for subtype A/H3. For the vaccine strain of the B/Yamagata lineage, B/Phuket/ 3073/2013 was selected, and B/Victoria/705/2018 (BVR-11) was selected as the vaccine strain for the B/Victoria lineage (https:// www.mhlw.go.jp/stf/shingi2/0000203023_00005.html).
A multicenter case-control study on the effectiveness of two doses of the inf luenza vaccine in children under three years of age was conducted, and two doses of vaccine were considered to reduce the risk of symptomatic inf luenza by about half (see p.255 of this issue).
Human infection with avian and swine inf luenza virus: Since September 2020, several human cases of subtype A/H5 inf luenza virus infection have been reported. Human cases of highly pathogenic avian inf luenza (HPAI) A (H5N1) virus infection have been reported from Laos and India, and human cases of A(H5N6) virus infection have been conf irmed in China and Laos. In December 2020, the f irst human case of A(H5N8) virus infection was also conf irmed in Russia. With the addition of these cases, the total number of human cases of inf luenza subtype A/H5 virus infection has reached 863 cases (including 456 fatal cases) since 2003. Human cases of A(H7N9) virus infection have been conf irmed in 1,568 cases (including 616 deaths) since 2013, but there have been no reports since March 2019. Human cases of A(H9N2) virus infection have been conf irmed in China and Cambodia since October 2020, but all cases were mild. In April 2021, the f irst human case of A(H10N3) virus infection was reported in China. In addition, HPAI due to subtype A/H5 viruses have occurred among poultry and wild birds in Asia, including Japan, Europe, and Africa, since September 2020.
Swine may be an intermediate “mixing vessel” host, generating reassortments of mammalian and avian inf luenza viruses. During the 2020/21 season, human cases of A(H3N2)v, A(H1N1)v, and A(H1N2)v virus infections were reported in the United States and Canada, and since October 2020, human cases of swine inf luenza viruses were also reported from countries such as Australia, China, Europe, and Brazil. Sporadic cases of human infection with avian inf luenza viruses have been reported in neighboring countries of Japan, and reassortant viruses of human A/H1pdm09 and swine inf luenza viruses have been detected in swine in Japan. It is necessary to monitor the occurrence of these viruses closely (see p.257 of this issue).
Conclusion: Since the 19th week of 2020, the number of reported inf luenza cases decreased and remained at a low level even during the regular inf luenza season in the 2020/21 season. Furthermore, there were no signs suggestive of an inf luenza epidemic based on multiple indicators. On the other hand, from the summer of 2021 onward, the number of reported inf luenza cases has been increasing overseas, suggesting the possibility that inf luenza activity may increase in the 2021/22 season (see p.259 of this issue). In preparation for the resurgence of inf luenza, it is important to continue public health measures such as vaccination of high-risk groups including the elderly, together with comprehensive monitoring of inf luenza, such as year-round isolation and detection of inf luenza viruses, analyses of the antigenic and genetic mutations of circulating strains, and surveys of the emergence of drug-resistant strains and seroprevalence levels in the population.
Note: For IASR reporting, inf luenza nomenclature is based on the virus information available. Inf luenza viruses are classif ied by type, subtype, lineage, and strain based on hemagglutination (HA), neuraminidase (NA), and other information including nucleotide sequence:
- When both HA and NA typing have been performed, the names are listed fully [e.g., A(H1N1)pdm09, A(H3N2), A(H5N1)].
- When NA typing has not been performed, only the HA information is listed (e.g., A/H1pdm09, A/H3).
- The strain name is represented by the location of isolation; in the case of isolation in Japan, the location is written in Japanese kanji, and in the case of isolation in foreign countries, in English.
- To distinguish swine inf luenza viruses recovered from humans from seasonal inf luenza viruses, the term “variant virus” is used with a letter “v” added after the subtype name [e.g., A(H3N2)v].
