高病原性鳥インフルエンザウイルスA(H5N1)感染事例に関するリスクアセスメントと対応
2023年4月13日
2025年4月1日最終更新
国立感染症研究所
更新点
2025年4月1日 疫学的所見、ウイルス学的所見、日本国内の対応、リスクアセスメント
目次
- 背景
- 疫学的所見
- 事例の概要
国外の状況
国内の状況 - 治療薬、ワクチン、検査について
- 事例の概要
- ウイルス学的所見
- 日本国内の対応
- リスクアセスメント
背景
高病原性鳥インフルエンザウイルスA(H5N1) (Highly pathogenic avian influenza virus: 以下、HPAIV(H5N1))は1997年に初めて、香港で生鳥市場を介したヒト感染例の報告があり、2003年、2004年には東アジア、東南アジアでもヒト感染例が報告された。これ以降、世界各地の家きんや野鳥に感染が拡がり、流行域を拡大したH5亜型のHPAIVは、A/goose/Guangdong/1/1996(H5N1)に由来するユーラシア型のヘマグルチニン(HA)遺伝子を保持しており、HA遺伝子の塩基配列により当初は0~9のCladeに分類され、その後HA遺伝子の変異が蓄積し、Cladeごとにさらに細かな亜系統に分類されるようになった。さらに他のA型インフルエンザウイルスとの遺伝子再集合を起こすなど、遺伝的にも多様化している。特に2005年以降はClade 2の亜系統が鳥類で流行したことに伴い鳥類からヒトへの感染例も増加し、2006年には欧州、アフリカ大陸でもヒト感染例が報告された。HPAIV(H5N1)のヒト感染例は2003年から2025年2月5日までで少なくとも965例が世界保健機関(WHO)に報告されており、2017年以前の報告が大部分を占めるが、2024年には北米からの報告数が増加した。
2021年以降はClade 2.3.4.4bのHPAIV(H5N1)の世界的な感染拡大に伴い、2023年には南極地域で初めて鳥類での感染例の発生が報告され、オセアニアを除く全世界から報告があったほか、水生動物を含む野生の哺乳類や農場のミンクなどの感染例、散発的なヒト感染例が世界各所で継続的に報告されている。加えて2024年3月には、米国からヤギおよび乳牛でのClade2.3.4.4bのHPAIV(H5N1)感染例、および未殺菌乳(生乳)からの同CladeのHPAIV検出が報告され、接触者の調査中にヒトの感染例が確認された。 また、Clade 2.3.2.1cのHPAIV(H5N1)の局地的なヒト感染例も報告されている。
近年のHPAIV(H5N1)のヒト感染例の報告は限られるが、鳥類や哺乳類で流行が拡大していることから、2020年以降の状況について、HPAIV(H5N1)感染事例の疫学情報の更新およびリスクアセスメントを行った。
疫学的所見
1.事例の概要
国外の状況
国外の鳥類(野鳥、家きん)における発生状況
Clade 2.3.4.4bの HPAIV(H5N1)は、2020年後半に欧州北部で同定されたのち、渡り鳥により世界各地へと広がった。鳥類における感染事例が確認された地域は、2021年から2022年にかけては欧州が主であり、北米にも拡大した(WHO. 2022)が、2023年には南米に広がり(OFFLU. 2023)、さらに2023年末から2024年2月にかけては南極大陸を含む南極地域にも拡大した(CSIC. 2024)。
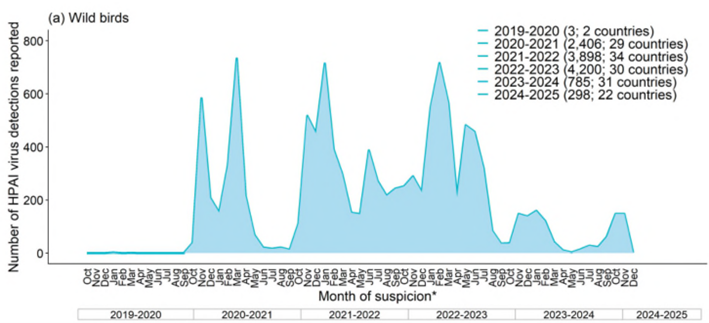
例年、鳥類におけるA型インフルエンザウイルス感染事例の報告数は、9月が最も少なく、10月頃から増加し始め、2月にピークを迎える(WOAH. 2023)。しかし、2021/2022シーズン※は例年報告数が減少する時期にも、欧米を主として報告数が減少しないままに2022/2023シーズンを迎えた。2021/2022および2022/2023シーズンの鳥類におけるHPAIV感染事例は例年にない規模となったが、2022/2023シーズンは例年と同様、7月から9月に報告数が減少した。2023/2024シーズンは、例年より早い1月にピークを迎え、その後減少し、過去3シーズンと比較すると報告数が少なかった(図1)(ECDC.2024d、WOAH. 2025)。
2024/2025シーズンは、10月以降現在も継続的に事例が発生しており、1月までの4か月間で報告された家きんの発生件数は、前シーズン通年と同程度の水準であった。現時点で、家きんにおいてはピークを過ぎている可能性がある。野鳥においては欧州での発生が増加している。(WOAH. 2025)。事例は主に欧米から報告されたが、日本を含むアジアからも報告されている(図2)。
※インフルエンザのシーズンの定義は地域などによって異なり、WHOなどは第40週(日本では第36週)から翌年の第39週(日本では第35週)までの1年間を1シーズンとしてカウントする。
(a)野鳥
(b)家きん
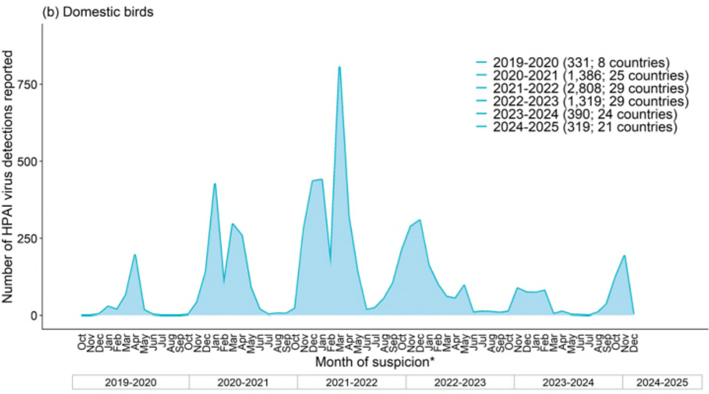
図1. 欧州における野鳥・家きんでのHPAIV検出状況
(2019年10月1日~2024年12月6日) (ECDC. 2024d)
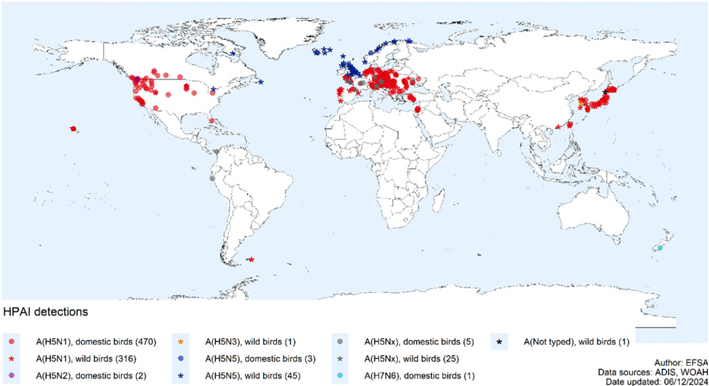
図2.鳥におけるHPAIV(H5N1)感染事例の報告状況
(2024年9月21日~12月6日) (ECDC. 2024d)
国外の哺乳類における発生状況
哺乳類におけるHPAIV(H5N1)感染事例は、2003年から確認されるようになり、主に、鳥類におけるアウトブレイク発生地で、野鳥を捕食することがある哺乳類を中心に発生していた(Plaza PI. et al.. 2024、CDC. 2023)。
ヒト以外の哺乳類におけるH5N1感染事例の発生国は、2003年から2019年までの17年間では10ヵ国に留まっていたが、2020 年から2023年10月までの約4年間で26ヵ国(欧州17ヵ国、南米5ヵ国、北米2ヵ国、アジア2ヵ国)と急増した(Plaza PI. et al.. 2024)。さらに、2023年12月には、南極地域においてもゾウアザラシのHPAIV感染が確認されており、哺乳類におけるHPAIV感染事例の発生地域は拡がっている(ECDC. 2024a、OFFLU. 2023)。
H5N1感染が確認された哺乳類は2020年から2023年10月までに48種以上と、多様な哺乳類における感染が確認されるようになった。2019年以前は陸生動物、半水生動物の感染が報告されていたが、2020年以降は水生動物(アザラシ、アシカ等)の感染も確認されるようになった(Plaza PI. et al.. 2024)。
哺乳類におけるH5N1感染事例は、野生動物に限らず飼育動物でも発生している。2022年10月にはスペインの大規模なミンク農場における複数のミンクの感染事例が報告された(Agüero M. et al.. 2023)。また、2023年にはフィンランドの複数の毛皮農場(ミンク、キツネ、タヌキ)における大規模感染等が報告された。このうちの多くは2023年9月から開始された後ろ向きの血清学的調査により探知された事例であり、死亡個体や有症個体が検出された農場は一部であった。このため、無症状で探知されていない事例があると考えられている(ECDC. 2024c)。
飼い猫(ポーランド、韓国、米国)、飼い犬(イタリア)における感染事例も報告されている(CDC. 2023)。中でも、2023年6月から7月にかけて、ポーランド国内複数地域から25匹の飼い猫のH5N1感染が確認された事例では、屋外での感染鳥類との接触だけでなく、エサとして与えられていた鶏肉も感染源の一つとして推察された(Domańska-Blicharz K. et al.. 2023)。また、2023年7月に発生した、韓国の2カ所の猫保護施設における数十匹の猫のH5N1感染事例では、市販のペットフードが感染源と考えられた(Kim. Y. et al.. 2023)。また、2024年12月には、米国オレゴン州およびネバダ州において、 2025年2月には、オレゴン州において、市販のペットフード、ならびに当該ペットフードを食べた猫から、H5N1(12月のオレゴン州)もしくはH5鳥インフルエンザ(ネバダ州および2月のオレゴン州)が検出されたことが報告された(County of Los Angeles、 Oregon Department of Agriculture)。
哺乳類における感染事例の多くは、単数もしくは少数個体の事例であるが、2022/2023シーズンには、アザラシ、アシカ等で数十頭から数百頭規模の大規模感染事例が報告され、哺乳類間での伝播が起きている可能性が示唆された(ECDC. 2024a、Puryear W. et al.. 2023)。また、南米のペルー、チリでは感染したアシカやゾウアザラシの大量死が報告されており、一部の地域で、その致命率の高さも懸念されている(OFFLU. 2023)。
米国のミネソタ州において、2024年3月20日、哺乳類の家畜では初となる、Clade2.3.4.4bのHPAIV(H5N1)のヤギでの感染事例が報告された(Minnesota Board of Animal Health, 2024)。3月25日にはカンザス州とテキサス州における乳牛の感染事例および未殺菌乳(生乳)からの同Cladeに属するHPAIV(H5N1)検出が報告された(USDA. 2024a、CDC. 2024b)。4月2日時点で5州(テキサス、カンザス、ミシガン、ニューメキシコ、アイダホ)において乳牛からのHPAIV(H5N1)検出の報告があり、テキサス州の事例では農場の調査中に農場内で死亡したネコや野鳥からもHPAIVが確認された(USDA. 2024b、Cornell University. 2024)。その後も、米国内の複数州から乳牛の感染事例の報告が相次ぎ、2025年2月6日時点で、16州において乳牛からのHPAIV(H5N1)検出が報告された(USDA. 2025a)。2024年3月以降に乳牛から検出されたClade 2.3.4.4bのHPAIV(H5N1)は、*遺伝子型B3.13であり、野鳥から乳牛へHPAIVのスピルオーバーが生じていたと考えられている(USDA. 2025b)。
(*遺伝子型: 同一CladeのHPAIV(H5N1)であっても、HA遺伝子以外のウイルス遺伝子の由来が異なる場合は、細分類された遺伝子型により区別する)
米国の乳牛における地域的な感染伝播の継続については、家畜の移動に加え、人、車両、その他の農機具の農場間での移動が要因と指摘されている(USDA. 2024d)。
2025年1月にネバダ州の酪農場の乳牛から採取された検体からは、Clade 2.3.4.4bのHPAIV(H5N1)、遺伝子型D1.1が検出された (USDA. 2025b、USDA. 2025c)。また、2025年2月にも、アリゾナ州の乳牛からClade 2.3.4.4bのHPAIV(H5N1)、遺伝子型D1.1が検出された(APHIS 2025)。遺伝子型D1.1は2024-2025年冬季の北米の野鳥において優勢であり、野鳥から乳牛へのHPAIV(H5N1)のスピルオーバーとして2・3例目の事例であると考えられている(USDA 2025b、APHIS 2025)。
また、米国のオレゴン州では、2024年10月30日、家きんでHPAIV(H5N1)が検出されていた非営利農場において、豚からHPAIV(H5N1)の検出が報告された(USDA. 2024e)。これは米国で初めての豚のHPAIV(H5N1)感染事例である。豚および同農場の家きんから検出されたClade 2.3.4.4b のHPAIV(H5N1)の遺伝子型はともにD1.2であり、この地域の渡り鳥のサンプルのゲノム配列と非常に類似した配列を示していた。このことから、この農場の豚や家きんは、乳牛や他の家畜ではなく、感染した渡り鳥との接触により感染した可能性が高いと考えられている (USDA. 2024f)。
世界的な哺乳類における感染拡大の背景には、H5N1ウイルスの遺伝的変異が関与している可能性が示唆されているが、現時点で、ヒトへの感染力が強まったとする報告はない(CDC. 2024d)。
国外のヒトにおける発生状況
WHOに報告されたヒトにおけるHPAIV(H5N1)感染事例は、2003年から2025年1月28日時点で合計966例あり、少なくとも466例(48%)が死亡している。このうち、2017年までの報告が860例(うち死亡454例(53%))と多くを占め、2018年以降の報告数は大きく減少していたが、2024年には報告数が増加した。HPAIV(H5N1)はヒト症例が報告されている鳥インフルエンザの中でも報告された症例数が多く、また致命率が高いウイルスである(表1)(WHO. 2024a、ECDC. 2024d、ECDC. 2025a、ECDC. 2025b)。
ヒト感染例は、2020年1月から2025年1月28日までに12ヵ国から106例が報告された(表2)。ほとんどの症例に、病気または病気の疑いがある動物や、死亡した家きんとの接触があった。このうち、ベトナム、カンボジア、オーストラリア以外の9ヵ国から報告されたHPAIV(H5N1)症例のうち、Cladeが確認できた症例はすべて2.3.4.4bであった。
鳥類および哺乳類におけるHPAIV(H5N1)感染事例の報告数が増加した2021/2022および2022/2023シーズンにも、ヒト感染例の著明な増加は確認されなかった。2023/2024シーズンおよび2024/2025シーズン(2023年9月以降から2025年2月時点)におけるヒト感染例は、ベトナム、中国、カンボジア、オーストラリア、米国、カナダ、英国から報告があったが、米国からの報告が最多であった。また、国外で報告されたヒト感染例の多くは感染した家きんや乳牛との接触歴があり、ヒト-ヒト感染を示唆する情報は確認されていない(CDC. 2024a、CDC. 2025、 WHO. 2025)。
2024年以降にHPAIV(H5N1)のヒト感染例が報告された国のうち、カンボジア、米国、オーストラリア、カナダ、英国での発生状況や事例の詳細について以下に示す。
カンボジアでは2014年から長期間、鳥類からのClade 2.3.2.1cの検出が続いているが、2015年から2022年まではヒト感染例は確認されていなかった。しかし、2023年以降、17例のヒト感染例が報告されており、情報のない症例を除き、検出されたHPAIV(H5N1)のCladeはすべて2.3.2.1cであった。
米国においては、2024年4月に乳牛に関連した症例が報告されて以降、2025年1月28日までに10州67例の HPAIV(H5)に感染したヒト症例が報告されている。感染源としては、23例はHPAIV(H5N1)に感染した家きんとの接触、40例は感染乳牛との接触、1例はその他動物との接触、3例は不明であった。多くの症例は軽症で、結膜炎や軽度な上気道症状を呈していた (CDC. 2024g)。2024年6月から8月に、ミシガン州とコロラド州の感染牛が確認された酪農場の従業員を対象とした血清学的調査によると、115人中8人(7%)でHPAIV(H5)に対する抗体を有していることが確認された(CDC. 2024h)。2024年12月には、ルイジアナ州において、高齢で基礎疾患があった患者で国内初の死亡例が報告された。患者は、米国で実施されている入院患者におけるインフルエンザサーベイランスにて探知された。遺伝子型はD1.1であり、米国内の野鳥や家きん、ならびにワシントン州およびカナダのヒト症例で検出されたウイルスと近縁であった。(CDC. 2024j)。
カナダにおいては、2024年11月にヒト感染例が報告され、HPAIV(H5N1)のCladeは2.3.4.4bであった。遺伝子型がD1.1であり、米国の乳牛で発生している遺伝子型B3.13とは違い、カナダ国内の家きんで発生しているHPAIV(H5N1)との関連が示唆された。しかし、症例における家きんとの接触は確認できていない(WHO. 2024c、WPRO. 2024、PHAC、PAHO. 2024)。
英国においては、2025年1月に2023年以来のHPAIV(H5N1)のヒト感染例が報告された。Cladeは2.3.4.4b、遺伝子型はDI.2であり、今シーズン英国での鳥で循環していることが知られており、米国の哺乳類や鳥類の間で循環している株とは異なる。(UKHSA. 2025、ECDC. 2025b)
オーストラリアからも、2024年5月に国内初のヒト感染例が報告された。検出されたHPAIV(H5N1)のCladeは2.3.2.1aであり、同一Cladeのウイルスが過去に鳥類で検出されているインドへの渡航歴があったことから、インドで曝露した可能性が考えられている(WHO. 2024b)。
2025年1月28日時点でH5N1以外のHPAIV(H5)のヒト感染例はH5N6で93例、H5N8で7例、H5N2で1例が報告されている(表1)(ECDC. 2024d)。また、H5以外の鳥インフルエンザでは低病原性鳥インフルエンザ(LPAIV)(H7N9)とHPAIV(H7N9)を合わせて1568例、HPAIV(H7N7)で94例、H9N2で151例のヒト感染例の報告があるほか、少数ではあるがH3N8、H6N1、H7N2、H7N3、H7N4、H10N3、H10N5、H10N7、H10N8の各亜型のヒト感染例が報告されている(WHO. 2019、WHO. 2023、WHO. 2024d、ECDC. 2024d、CDC. 2024e、Belser J. et al.. 2009、 Puzelli S. et al.. 2013)。
表1.鳥インフルエンザウイルス(H5亜型)のヒト感染事例(2025年1月28日時点)
|
亜型 報告年 |
H5N1 | H5N6 | H5N8 | H5N2 |
|---|---|---|---|---|
| -2009 | 468 | 0 | 0 | 0 |
| 2010-2014 | 233 | 3 | 0 | 0 |
| 2015 | 145 | 5 | 0 | 0 |
| 2016 | 10 | 9 | 0 | 0 |
| 2017 | 4 | 2 | 0 | 0 |
| 2018 | 0 | 4 | 0 | 0 |
| 2019 | 1 | 1 | 0 | 0 |
| 2020 | 1 | 5 | 7 | 0 |
| 2021 | 2 | 37 | 0 | 0 |
| 2022 | 6 | 18 | 0 | 0 |
| 2023 | 12 | 6 | 0 | 0 |
| 2024 | 81 | 3 | 0 | 2 |
| 2025 | 2 | 0 | 0 | 0 |
| 合計 | 965 | 93 | 7 | 2 |
(WHO. 2024d、WHO. 2025、ECDC. 2024b、ECDC. 2024d、ECDC, 2025a、ECDC. 2025b、UKHSA. 2025)
表2. 2020年以降に報告されたHPAIV(H5N1)ヒト感染事例(2025年1月28日時点)
| 報告年 | 報告国 | 報告数 | 死亡例数 | Clade |
|---|---|---|---|---|
|
2020 |
ラオス | 1 | 0 | 2.3.4.4b |
| 2021 | インド | 1 | 1 | 2.3.4.4b |
| 英国 | 1 | 0 | 2.3.4.4b | |
| 2022 | 中国 | 1 | 1 | 2.3.4.4b |
| エクアドル | 1 | 0 | 2.3.4.4b | |
| スペイン | 2 | 0 | 2.3.4.4b | |
| 米国 | 1 | 0 | 2.3.4.4b | |
| ベトナム | 1 | 0 | 情報なし | |
| 2023 | カンボジア | 6 | 4 | 2.3.2.1c |
| 中国 | 1 | 情報なし | 2.3.4.4b | |
| チリ | 1 | 0 | 2.3.4.4b | |
| 英国 | 4 | 0 | 2.3.4.4b | |
| 2024 |
カンボジア | 10 | 2 | 5例:2.3.2.1c 5例:情報なし |
| ベトナム | 1 | 1 | 2.3.2.1c | |
| 米国 | 67 | 1 | 2.3.4.4b* | |
| オーストラリア | 1 | 0 | 2.3.2.1a | |
| 中国 | 1 | 0 | - | |
| カナダ | 1 | 0 | 2.3.4.4b | |
| 2025 | 英国 | 1 | 0 | 2.3.4.4b |
| カンボジア | 1 | 1 | 2.3.2.1c |
* GISAIDのEpiFluTM データベースには2.3.4.4bに属するHPAIVのゲノムデータが55件登録されており、他クレードは登録されていない
(WHO. 2024a、WHO. 2024d、WHO. 2025a、WHO. 2025b、CDC.2024b、ECDC. 2024a、ECDC. 2024b、ECDC. 2024d、ECDC. 2025a、ECDC. 2025b、PHAC、UKHSA. 2025、GISAID. 2025)
国内の状況
国内の鳥類における発生状況
2023/2024シーズンに国内で得られた全11例のHPAIVのHA遺伝子は、Clade 2.3.4.4bに属 しており、2017/2018、2020/2021、2021/2022および2022/2023シーズンと同じ系統であった(農林水産省. 2024a)。
2024/2025シーズンは、野鳥、家きん、それぞれ、2024年9月30 日と10月17日に国内1事例目が確認され、2023/2024シーズンと比較すると1事例目が確認された時期が早かった(農林水産省. 2024b、2025)。
2024/2025シーズンの国内の鳥におけるHPAIV感染事例は、2025年2月5日時点で、野鳥では17道県から112事例(うち、H5N1は17道県109事例)が、家きんでは14道県から51事例(うちH5N1は14道県47事例)が報告され、約932万羽が殺処分対象となった。飼養鳥においては、2025年2月5日時点での報告はない(環境省. 2025、農林水産省.2025)。
2024/2025シーズンの家きんにおける発生件数(2025年2月1日時点で14道県51事例発生)は、過去最多の発生となった2022/2023シーズン(同日時点で25道県71事例発生)に比べ、同日比で7割強となった(農林水産省.2025)。
国内の哺乳類における発生状況
2022年4月に北海道札幌市において、キタキツネ(アカギツネ)およびタヌキでのHPAIV(H5N1)感染事例が国内で初めて確認された(磯田ら. 2022)。2023年4月と6月には同市において2例のキツネでのH5N1感染(死亡個体からの検出、6月探知例の検体採取月は同年2月)が探知された。周辺地域ではハシブトカラスのHPAIV(H5N1)感染事例が続発しており、キタキツネおよびタヌキに感染していたHPAIV(H5N1)は、ハシブトガラスから検出されたウイルスと遺伝的に類似していた(Hiono T. et al.. 2023)。キタキツネについては、HPAIV(H5N1)に感染した野鳥を捕食してHPAIV(H5N1)に感染した事が死因と考えられた。タヌキについては、他の病原体による感染も認められ、HPAIV(H5N1)感染が直接の死因か不明であった(磯田ら. 2022)。また、2023/2024シーズンにおけるHPAIの発生に係る疫学調査では、11事例中4事例で家きん舎内の環境材料を用いた検査において検体からHPAIVが検出されている。このうち、広島県の発生事例では、発生鶏舎の隣の鶏舎で死亡していたクマネズミからHPAIVが検出されており、これは対応する発生農場の家きん由来HPAIV(H5N1)と同一遺伝子型で、対応する農場由来ウイルスとの間で99パーセントを超える極めて高い一致率を示した(農林水産省. 2024b)。
2025年2月5日現在、今シーズンは哺乳類においてHPAIVは確認されていない。
国内のヒトにおける発生状況
国内ではこれまでにHPAIV(H5N1)を含め、鳥インフルエンザウイルスに感染して発症したヒト感染事例は確認されていない。
2.治療薬、ワクチン、検査について
抗インフルエンザ薬、特にノイラミニダーゼ(NA)阻害薬やポリメラーゼ阻害薬に対する耐性を獲得しているHPAIV(H5N1)の流行は認められていないため、これらの薬剤による治療効果は期待できる。
近年、ヒトおよび哺乳動物で感染が確認されているClade 2.3.4.4bのHPAIV(H5)ウイルスについては、WHOより以下が使用可能なワクチン候補株としてリストされている(WHO 2025a, WHO 2025b)。A/Astrakhan/3212/2020(IDCDC-RG71AおよびCBER-RG8A, H5N8) A/American wigeon/South Carolina/22-000345 -001/2021(IDCDC-RG78, H5N1)、A/Ezo red fox/Hokkaido/1/2022 (NIID-002, H5N1)、A/Jiangsu/NJ210/2023(CNIC-JSNJ210, H5N1)。また近年、ヒト感染が確認されたClade 2.3.2.1cのH5ウイルスについては、A/duck/Vietnam/NCVD-1584/2012(NIBRG-301, H5N1)が使用可能なワクチン候補株として上記リストに掲載されている。
HPAIV(H5N1)を含むA型インフルエンザウイルスの検出に関しては、呼吸器検体を用いたコンベンショナルRT-PCRもしくはリアルタイムRT-PCR法によるウイルス遺伝子検出検査の実施が推奨されている。検査に使用する検体は鼻腔スワブ(鼻の奥)、口腔咽頭スワブ(喉)、鼻咽頭スワブ(鼻咽頭)に加え、鼻咽頭吸引液や気管支吸引液などが有用とされている(WHO. 2021)。
ウイルス学的所見
Clade 2.3.4.4bのHPAIV(H5N1)は、2020年後半に欧州北部で同定された後、渡り鳥により世界各地へと運ばれ、様々な国・地域で遺伝子再集合(他のA型インフルエンザウイルスとの遺伝子分節の交換)した多様な遺伝子型のHPAIV(H5N1)が分離されている(Leguia M. et al.. 2023、Alkie TN. et al.. 2023)。ただし、遺伝子型の違いによるウイルス性状の違いはよく分かっていない。
鳥類から分離されたClade 2.3.4.4bのHPAIV(H5N1)からは、哺乳類での病原性や増殖能力の獲得に寄与するPB2タンパク質のE627K変異を持つウイルスや、HAタンパク質の受容体結合部位にヒト型受容体(α2,6結合したシアル酸)への結合能力の増強の可能性が示唆されるアミノ酸変異を持つウイルス(例えば、S137A, T160Aなど)等がまれに報告されている(ECDC. 2024d)。
2022年10月にスペインのミンク農場のミンクから分離されたClade 2.3.4.4bのHPAIV(H5N1)には、哺乳類由来細胞内でのポリメラーゼ活性の上昇に関与するPB2タンパク質のT271A変異が認められた(Agüero M. et al. 2023)。また、2022年4月から7月にかけてカナダの野生のアカギツネ、スカンク、ミンクから分離されたClade 2.3.4.4bのHPAIV(H5N1)の40株全てのウイルスのHAタンパク質にS137AおよびT160A変異が認められ、そのうちの17%は、哺乳類への適応に関与するPB2のE627K、E627V、D701Nのいずれかのアミノ酸変異が認められた(Alkie TN. et al. 2023)。
2024年3月に米国テキサス州の乳牛でHPAIV(H5N1)感染例が世界で初めて報告された。感染牛の口腔咽頭ぬぐい液、生乳からClade 2.3.4.4b(遺伝子型B3.13)のHPAIV(H5N1)が検出された(Burrough et al.. 2024)。また、感染牛と同じ農場内で死亡したネコと野鳥からも類似したゲノム配列を有するHPAIV(H5N1)が検出され、ネコへの感染は病牛の初乳を与えられたことによる水平伝播であることが示唆された。哺乳類間の水平伝播は、既報のネコ(Kuiken T et al.. 2004)およびフェレット(Herfst S et al.. 2012)への感染実験でも証明されている(Burrough et al.. 2024)。
生乳およびネコから検出されたHPAIV(H5N1) (遺伝子型B3.13) と過去に陸生および海生哺乳類から検出されたウイルスの遺伝子配列を比較したところ、生乳およびネコから検出されたウイルスではHAタンパク質内にヒト型レセプターとの結合親和性を高めると考えられているアミノ酸残基S137A、N158N、T160Aの各アミノ酸変異が確認されたが、T192I、G225D、G228Sを含むものはなかった(Yamada S et al.. 2006, Gao Y, et al.. 2009, CDC. 2024d)。これらのアミノ酸変異(S137A、N158N、T160A)は過去の陸生および海生哺乳類から分離されたHPAIVで多く観察されたものと同一であった。以前から哺乳類での病原性や増殖能力の獲得に寄与することが知られているPB2のアミノ酸変異T271A、I292V、Q591K、E627K/V/A、D701Nは、当該分離株すべてには見られなかったが、テキサス州における乳牛からの感染が推定された最初のヒトの症例から分離されたHPAIVはPB2にE627 K変異を示した(Gao Y, et al.. 2009, Suttie A et al.. 2019, Bordes L et al.. 2023, Hatta M et al.. 2007, Kong H et al.. 2019, Hu et al.. 2024)。テキサス州からの生乳およびネコ由来8株のHPAIVとヒト事例から検出されたHPAI (H5N1)ウイルスの遺伝子配列には、いずれも、現在、米国食品医薬品局(U.S. Food and Drug Administration: FDA)が推奨している抗ウイルス薬に対する感受性を低下させる既知のアミノ酸変異は含まれていなかった(Hu et al.. 2024, CDC. 2024i)。
哺乳類で分離されたウイルスのゲノム配列の定期的なモニタリングとスクリーニングでは、Clade2.3.4.4bのHPAIV(H5N1)に哺乳類適応のアミノ酸変異はほとんど見つかっておらず、2025年12月20日現在、米国の乳牛でウイルスの循環が続いているにもかかわらず、HA遺伝子の受容体結合能の変化に関連するアミノ酸変異は確認されていない(WHO. 2024e)。
コロラド州の養鶏場従業員のヒト事例から検出されたウイルスは、Clade 2.3.4.4bのHPAIV(H5N1)であり、ミシガン州の病牛がいる農場労働者の症例から検出されたウイルスと類似していた(CDC. 2024i)。この配列は主に鳥類に感染するHPAIVの遺伝的特徴を維持しており、ウイルスをヒトに感染させたり、ヒトの間で拡散させたりするような適応性を高めるような変化は見られない。また、PB2のM631L変異があり、これは2024年4月までに乳牛から検出されたウイルスの99%で確認されている哺乳類への適応に関わるアミノ酸変異と同じもので、ミシガン州の最初のヒト感染例でも確認された(CDC, 2024i)。一方、この配列にはテキサス州の症例のウイルスに見られたPB2 E627K変異は確認されなかった(CDC. 2024g)。
2024年12月13日にCDCから報告されたルイジアナ州初のヒト感染例で検出された HPAIV(H5N1)は、D1.1遺伝子型と特定された(CDC.2024j)。患者口腔咽頭スワブを含む検体中のウイルスのHAに、ヒト型レセプターへの結合に関与する変異(A134V、 N182K、E186D)が低頻度で認められた。これらの変異は、ヒトから分離された HPAIV(H5N1) ではまれであるが、これまでにも他国での過去のHPAIV(H5N1)症例で報告されており、たいていは重症感染例で見られるものである。このうち、E186Dは、患者内で複製される過程で出現したことが示唆されており、カナダのブリティッシュコロンビア州で重症のヒト症例から採取された検体中のウイルスでも確認されている(PHAC. 2024)。なお、NA、M、PA遺伝子に抗ウイルス薬に対する感受性を低下させる既知の、あるいは疑われるマーカーに関連する変化は見られなかった(CDC, 2024j)。
2025年1月28日にUKHSAから報告された養鶏場従業員1例から検出されたウイルスは、英国の鳥類で流行しているウイルスであるDI.2遺伝子型であり、PB2のI292V変異があった。しかしながら、当該変異は、鳥類から分離された株でもよく見られるものである(ECDC. 2025b)。
テキサス州で発生したヒトの症例に由来する B3.13 ウイルスを含む Clade 2.3.4.4bのHPAIV(H5N1)を用いたフェレットの感染実験では、直接接触によるフェレット間の伝播は様々であるが、呼吸器飛沫を介した伝播はほとんど見られないか、非効率的であることが示されている(CDC. 2024k,Pulit-Penaloza et al.. 2024, Eisfeld AJ et al.. 2024,Restori KH et al.. 2024,Pulit-Penaloza JA et al.. 2024)。 また、遺伝子型B3.13ではないが、チリで重篤な症状を示したヒトから分離されたClade2.3.4.4bのHPAIV(H5N1)は、眼球に感染させたフェレットに重篤な疾患を引き起こし、直接接触によって他のフェレットへもウイルス感染を起こすことができた(Belser JA et al.. 2024)。
オハイオ州で牛から分離されたClade 2.3.4.4bの HPAIV(H5N1) (遺伝子型B3.13)をカニクイザル(ヒト感染モデル)の鼻腔内または気管内に接種した動物実験では、それぞれ軽度および重度の呼吸器疾患を引き起こした。また、経口接種では感染は限定的であり、不顕性感染にとどまることが明らかになった (Rosenke K et al.. 2025)。
上述した鳥類や哺乳類から分離されたClade 2.3.4.4bのHPAIV(H5N1)に認められる、哺乳類適応やヒト型受容体への結合能に関与する可能性のあるアミノ酸変異によるヒト感染への直接的な影響についてはよく分かっていない。現在までのところ、Clade 2.3.4.4bのHPAIV(H5N1)の効率的なヒトーヒト感染は報告されておらず、ウイルス学的および疫学情報からは、これらのウイルスが哺乳類宿主への適応を確立しておらず、ヒトからヒトに持続的に感染する能力を獲得しているという証拠もない(WHO. 2024f)。
カンボジアでは2023年に6例、2024年に10例のHPAIV(H5N1)(2025年1月28日現在)の感染が報告された(うち、死亡例は6例)。このうち、遺伝子解析結果が判明しているウイルスのHA遺伝子は、Clade 2.3.2.1cに属していた(WPRO. 2024)。Clade 2.3.2.1cのHPAIV(H5N1)は、2020年以降地理的に限定されたアジア地域の家きんで限局的に報告されている(GISAID. 2024)。Clade 2.3.2.1cのHPAIV(H5N1)についても持続的なヒトーヒト感染は報告されていない。
日本国内の対応
1. 国内における鳥インフルエンザウイルスのヒト感染事例の探知と対応について
鳥インフルエンザ(H5N1)は、「感染症の予防および感染症の患者に対する医療に関する法律」(感染症法)で定める二類感染症の「特定鳥インフルエンザ」の一つとして政令で指定されており、医師は鳥インフルエンザ(H5N1)の患者、無症状病原体保有者、疑似症患者を診断したとき、また、感染症死亡者の死体、感染症死亡疑い者の死体を検案したときは、感染症法第12条に基づき症例を届け出なければならない。都道府県知事等が感染症法第15条に基づき積極的疫学調査を実施する場合については、特に重症の患者の見逃しを防ぐために「鳥インフルエンザ(H5N1)に関する積極的疫学調査の実施等について(依頼)」(令和6年12月12日付け感感発1212第1号厚生労働省健康・生活衛生局感染症対策部感染症対策課長通知)において、その目的、実施体制を含めた技術的助言がなされている。この中では、近年の鳥インフルエンザ(H5N1)の発生状況に基づき、感染したヒト、鳥類に加え、哺乳類との接触がある場合、排せつ物や未殺菌の肉、生乳等との接触がある場合についても要観察例として積極的疫学調査の対象とすることを推奨している。
動物への職業曝露等が避けられない場合には、国内各省庁、各機関からの注意喚起や対策に従い、適切な対処を行う必要がある(農林水産省2024d、環境省2024)。
2. 国内における鳥インフルエンザウイルスの動物感染事例の探知と対応について
獣医師又は感染鳥類の所有者は、鳥インフルエンザ(H5N1)に感染している、若しくはその疑いのある鳥類を認めた場合は、感染症法第13条に基づき届け出なければならない。届出を受けた都道府県知事等は、感染症法第15条に基づく調査および法第29条に基づく措置等を行う。この際の対応については「国内の鳥類における鳥インフルエンザ(H5N1)発生時の調査等について」(平成18年12月27日付け健感発第1227003号厚生労働省健康局結核感染症課長通知(令和5年11月 10日一部改正))に基づき実施する(厚生労働省. 2006)。
2024年10月以降、国内複数地域での鳥類における感染事例の発生を受け、環境省ではレベル3(国内複数個所や近隣諸国での発生時)の対応として、鳥類生息状況等調査による監視強化、死亡野鳥等を対象にしたウイルス保有状況調査を強化している。
2024/2025シーズンは2024年10月18日に開催された第91回家きん疾病小委員会において、全国各地で環境中ウイルスが増加し、発生リスクが高まっていることから、基本的な飼養衛生管理の徹底等を含む今後の防疫対応の徹底について、取りまとめを行った。さらに2024年11月21日には鳥インフルエンザ防疫対策緊急全国会議を開催し、今後の対策強化の4つのポイント 1飼養衛生管理の「隙」を埋める対策、2再発対策(既発農場・地域への指導強化)、3大規模農場対策、4発生時の速やかな防疫措置、について発信した。2024年12月末までの発生件数は2024年10月17日の発生に始まり計16件にとどまっていたものの、2025年1月には愛知県、千葉県および岩手県の特に養鶏密集地域における続発が相次ぎ、1月2日から2月1日までの約1か月間で35件の発生が確認された。これは1か月間に確認された発生数としては過去最大の規模となった。このような発生状況を踏まえて、2025年1月20日には第2回の緊急全国会議を開催し、改めて危機感を共有するとともに、愛知県の事例においてまん延防止対策としての効果が示唆された鶏舎の入気・塵埃対策としての不織布シートや液状消毒薬の活用についても情報発信した。また、当該3県については、農林水産省の現地対策本部を設置し、県と一体となったまん延防止対策を推進した。
また、米国の乳牛におけるHPAIV(H5N1)感染例が確認されたことをうけ、農林水産省は2024年4月3日に都道府県や畜産関係団体に対し、米国の事例の共有および国内飼養牛において乳量の減少や食欲低下等がみられた際の対応、HPAI等の野鳥からの感染防止を図るための飼養衛生管理の徹底について、通知を発出した (農林水産省.2024c)。
各国・各機関におけるリスクアセスメント
2020年以降のClade 2.3.4.4b のHPAIV(H5N1)によるヒト感染事例と鳥類および哺乳類の感染事例の報告が増加したことにより、 WHO、欧州疾病予防対策センター(ECDC)、 CDC、英国健康管理庁(UKHSA)は、以下の通りリスクアセスメントを発出している(表3)。一方、Clade 2.3.2.1cのHPAIV(H5N1)は2020年以降についてはアジアで限局的に循環しており、世界的な感染拡大はみられていない。
CDCは動物で循環しているA型インフルエンザに対するパンデミックポテンシャルのリスク評価を行い、パンデミックに備えるべきウイルスの優先順位を決定するためにInfluenza Risk Assessment Tool (IRAT)を提唱している。本ツールには、ヒトーヒト感染持続の可能性(emergence)とヒトーヒト感染が持続した際の公衆衛生へのインパクト(public health impact)という2つの評価分野があり、それぞれについてリスク評価が行われる(国立感染症研究所. 2015)。H5N1ウイルスは主要なClade、株ごとに評価されており、Clade 2.3.4.4bのHPAIV(H5N1)は、中レベルのリスクに分類されている(CDC. 2024a)。
また、FDAと米国農務局 (U.S. Department of Agriculture: USDA)は、2024年の米国におけるヤギ、乳牛のHPAIV感染例の発生および未殺菌乳(生乳)からのHPAIV検出を受けて、国内で流通している牛乳の安全性には影響がないとの声明を発表している (USDA. 2024c)ほか、CDCも一般市民がHPAIV(H5N1)に感染するリスクは依然として低い、としている (CDC. 2024c)。
国立感染症研究所におけるリスクアセスメントと推奨
【海外渡航者が感染するリスク】
- 海外でのヒト感染例の多くは感染した家きん類等との接触による散発的な感染であり、効率的なヒトーヒト感染を示唆する情報はないことから、鳥類への曝露機会がない海外渡航者が感染する可能性は低い。
- 海外渡航者は、家きん市場や生きた鳥類、鳥類や哺乳類の死骸に不用意に近づかないように注意すべきである。
- 発生地域において鳥類との接触があり、渡航後に発熱を認めるなどの体調の変化があった場合には、医療機関の受診時に渡航歴および鳥類との接触歴を伝えることの啓発が必要である。
【国内で鳥類、哺乳類への接触者が感染するリスク】
- これまで国内で明らかなヒト感染例の報告はなく、ヒトへの感染性が高くなったという証拠は無いことから、鳥類への曝露機会がない人々への感染リスクは低い。一方、国内でも鳥類でのHPAIV(H5N1)検出事例が継続して報告されていることから、生きた鳥類や鳥類の死骸に不用意に近づかないように注意すべきである。
- 同様に哺乳類からヒトが感染するリスクも低いものの、国外で哺乳類の感染例の報告が増加していること、国内でも限定的ながら哺乳類での検出事例の報告があることから、哺乳類の死骸にも不用意に近づかないように注意すべきである。
【HPAIV(H5N1)がヒトへの感染性を獲得するリスク】
- HPAIV(H5N1)について、哺乳類への適応やヒトへの感染性が高くなるウイルス学的性質の獲得に関する証拠は限定的であり、疫学的にも効率的なヒトーヒト感染の証拠はない。ただし、動物で感染が拡大する中でアミノ酸変異が蓄積して、ヒトへの感染性がより高くなったウイルスが今後出現する可能性は否定できないことから、引き続き動物での発生動向を監視する必要がある。
【HPAIV(H5N1)がヒトでパンデミックを引き起こすリスク】
- HPAIV(H5N1)は効率的にヒトからヒトへ感染する能力を獲得しておらず、現時点ではヒトでのパンデミックに至る可能性は低い。しかし、世界的に鳥類での感染拡大が認められ、哺乳類の感染例も多数報告されていることから、HPAIV(H5N1)へのヒトの曝露機会が増加しており、今後も散発的なヒト感染例が報告される可能性は高い。
- HPAIV(H5N1)に感染した鳥類や哺乳類とヒトとの接触頻度や感染リスク、そこからウイルスが効率的にヒトーヒト感染する能力を獲得するリスクを定量的に見積もるには十分な知見がないが、今後も感染動物とヒトとの接触機会を極力避けつつ、継続して動物での発生動向を監視し、適時にリスク評価を行う必要がある。
表3. WHO、ECDC、CDC、UKHSAによるHPAIV(H5N1)に関する状況のまとめとリスクアセスメント
| WHO | ECDC | CDC | UKHSA | |
|---|---|---|---|---|
| 状況のまとめ |
|
|
|
|
| リスクアセスメント |
|
|
|
|
(WHO. 2024b、ECDC. 2024d、CDC. 2024b、UKHSA. 2024)
関連項目
ポーランド共和国および大韓民国におけるネコの高病原性鳥インフルエンザウイルスA(H5N1)感染事例について
鳥インフルエンザA(H7N9)ウイルスによる感染事例に関するリスクアセスメントと対応
参考文献
- Agüero M, Monne I, Sánchez A, et al. Highly pathogenic avian influenza A(H5N1) virus infection in farmed minks, Spain, October 2022. Euro Surveill. 2023;28(3):2300001. doi:10.2807/1560-7917.ES.2023.28.3.2300001
- Alkie TN, Cox S, Embury-Hyatt C, et al. Characterization of neurotropic HPAI H5N1 viruses with novel genome constellations and mammalian adaptive mutations in free-living mesocarnivores in Canada. Emerg Microbes Infect. 2023;12(1):2186608. doi:10.1080/22221751.2023.2186608
- APHIS Identifies Third HPAI Spillover in Dairy Cattle. As of February 13, 2025,
- Belser JA, Bridges CB, Katz JM, Tumpey TM. Past, present, and possible future human infection with influenza virus A subtype H7. Emerg Infect Dis. 2009;15(6):859-865. doi:10.3201/eid1506.090072
- Belser JA, Sun X, Pulit-Penaloza JA, Maines TR. Fatal Infection in Ferrets after Ocular Inoculation with Highly Pathogenic Avian Influenza A(H5N1) Virus. Emerg Infect Dis. 2024;30(7):1484-1487.
- Bordes L, Vreman S, Heutink R, et al. Highly Pathogenic Avian Influenza H5N1 Virus Infections in Wild Red Foxes (Vulpes vulpes) Show Neurotropism and Adaptive Virus Mutations. Microbiol Spectr. 2023;11(1):e0286722. doi:10.1128/spectrum.02867-22
- Burrough ER, Magstadt DR, Petersen B, et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerging Infectious Diseases. 2024;30(7):1335-1343. doi:10.3201/eid3007.240508.
- CDC. Technical Report: December 2023 Highly Pathogenic Avian Influenza A(H5N1) Viruses. https://www.cdc.gov/bird-flu/php/technical-report/h5n1-122923.html 2023
- CDC. Results of Influenza Risk Assessment Tool 2024a.
- CDC. A(H5N1) Bird Flu Response Update November 4, 2024 2024b.
- CDC. Highly Pathogenic Avian Influenza A (H5N1) Virus Infection Reported in a Person in the U.S. released 1 April 2024. 2024c.
- CDC. Technical Report: June 2024 Highly Pathogenic Avian Influenza A(H5N1) Viruses. Updated June 5, 2024. 2024d.
- CDC. Reported Human Infections with Avian Influenza A Virus. As of 9 April 2024. 2024e.
- CDC. A(H5N1) Bird Flu Response Update November 18, 2024. 2024g.
- CDC. Serologic Evidence of Recent Infection with Highly Pathogenic Avian Influenza A(H5) Virus Among Dairy Workers ― Michigan and Colorado, June–August 2024. 2024h.
- CDC. Technical Report: June 2024 Highly Pathogenic Avian Influenza A(H5N1) Viruses. 2024. 2024i
- CDC. Genetic Sequences of Highly Pathogenic Avian Influenza A(H5N1) Viruses Identified in a Person in Louisiana. 2024j
- CDC. CDC Reports A(H5N1) Ferret Study Results. 7 June 2024. 2024k
- CDC. H5 Bird Flu Response: Focus Areas for Ongoing Public Health Risk Assessment. 2025
- Cornell University. Cornell scientists identify bird flu infecting dairy cows. released 2 April 2024.
- County of Los Angeles. Public Health Warns Against Feeding Pets Raw Food Following H5 Bird Flu Virus Detection. released 31 December 2024.
- CSIC (Consejo Superior de Investigaciones Científicas). Científicos del Centro de Biología Molecular Severo Ochoa del CSIC confirman la presencia por primera vez en la Antártida del virus de la Gripe Aviar Altamente Patogénica. Published on 25 February 2024.
- Domańska-Blicharz K, Świętoń E, Świątalska A, et al. Outbreak of highly pathogenic avian influenza A(H5N1) clade 2.3.4.4b virus in cats, Poland, June to July 2023. Euro Surveill. 2024 Mar;29(10). doi:10.2807/1560-7917.ES.2023.28.31.2300366. . 2023;28(31):2300366.
- ECDC. Avian influenza overview June–September 2024. 2024a.
- ECDC. Communicable disease threats report, 9-15 November 2024, week 46. 2024b.
- ECDC. Avian influenza overview December 2023–March 2024. 2024c.
- ECDC. Avian influenza overview September–December 2024. 2024d.
- ECDC. Communicable disease threats report, 24-31 January 2025, week 5 2025a
- ECDC Weekly Bulletin. 2025b
- Eisfeld AJ, Biswas A, Guan L, et al. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature. 2024;633(8029):426-432. doi:10.1038/s41586-024-07766-6
- Gao Y, Zhang Y, Shinya K, et al. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog. 2009;5(12):e1000709. doi:10.1371/journal.ppat.1000709
- GISAID. https://gisaid.org/ As of 13 February 2024. 2024
- GISAID. https://gisaid.org/ AS of 4 March 2025. 2025
- Hatta M, Hatta Y, Kim JH, et al. Growth of H5N1 influenza A viruses in the upper respiratory tracts of mice. PLoS Pathog. 2007;3(10):1374-1379. doi:10.1371/journal.ppat.0030133
- Herfst S, Schrauwen EJ, Linster M, et al. Airborne transmission of influenza A/H5N1 virus between ferrets. Science. 2012;336(6088):1534-1541. doi:10.1126/science.1213362
- Hiono T, Kobayashi D, Kobayashi A, et al. Virological, pathological, and glycovirological investigations of an Ezo red fox and a tanuki naturally infected with H5N1 high pathogenicity avian influenza viruses in Hokkaido, Japan. Virology. 2023;578:35-44. doi:10.1016/j.virol.2022.11.008
- Hu X, Saxena A, Magstadt DR, et al. Genomic characterization of highly pathogenic avian influenza A H5N1 virus newly emerged in dairy cattle. Emerg Microbes Infect. 2024;13(1):2380421. doi:10.1080/22221751.2024.2380421
- Kong H, Ma S, Wang J, et al. Identification of Key Amino Acids in the PB2 and M1 Proteins of H7N9 Influenza Virus That Affect Its Transmission in Guinea Pigs. J Virol. 2019;94(1):e01180-19. Published 2019 Dec 12. doi:10.1128/JVI.01180-19
- Kim Y, Fournié G, Métras R, et al. Lessons for cross-species viral transmission surveillance from highly pathogenic avian influenza Korean cat shelter outbreaks. Nat Commun. 2023;14(1):6958. Published 2023 Oct 31. doi:10.1038/s41467-023-42738-w
- Kuiken T, Rimmelzwaan G, van Riel D, et al. Avian H5N1 influenza in cats. Science. 2004;306(5694):241. doi:10.1126/science.1102287
- Leguia M, Garcia-Glaessner A, Muñoz-Saavedra B, et al. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat Commun. 2023;14(1):5489. Published 2023 Sep 7. doi:10.1038/s41467-023-41182-0
- Minnesota Board of Animal Health, Stevens County goat tests positive for same influenza virus affecting poultry. Released 20 March 2024.
- OFFLU. Continued expansion of high pathogenicity avian influenza H5 in wildlife in South America and incursion into the Antarctic region. 21 December 2023.
- Oregon Department of Agriculture. Northwest Naturals of Portland Voluntary Recall of Northwest Naturals Brand 2lb Feline Turkey Recipe Raw & Frozen Pet Food Due to HPAI Contamination. released 26 December 2024. https://apps.oregon.gov/oregon-newsroom/OR/ODA/Posts/Post/morasch-meats-voluntary-recall-feline-raw-pet-food-hpai
- Oregon Department of Agriculture. Detections of HPAI in Domestic Cats. released 14 February 2025. https://apps.oregon.gov/oregon-newsroom/OR/ODA/Posts/Post/detections-of-hpai-in-domestic-cats
- PAHO. Epidemiological Update Avian Influenza A(H5N1) in the Americas Region - 15 November 2024. https://www.paho.org/en/documents/epidemiological-update-avian-influenza-ah5n1-americas-region-15-november-2024/.
- PHAC. Public Health Agency of Canada (PHAC). Statement from the Public Health Agency of Canada: Update on Avian Influenza and Risk to Canadians. https://www.canada.ca/en/public-health/news/2024/11/update-on-avian-influenza-and-risk-to-canadians.html.
- Plaza PI, Gamarra-Toledo V, Euguí JR, Lambertucci SA. Recent Changes in Patterns of Mammal Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus Worldwide. Emerg Infect Dis. 2024;30(3):444-452. doi:10.3201/eid3003.231098
- Puryear W, Sawatzki K, Hill N, et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in New England Seals, United States. Emerg Infect Dis. 2023;29(4):786-791. doi:10.3201/eid2904.221538
- Pulit-Penaloza JA, Brock N, Belser JA, et al. Highly pathogenic avian influenza A(H5N1) virus of clade 2.3.4.4b isolated from a human case in Chile causes fatal disease and transmits between co-housed ferrets. Emerg Microbes Infect. 2024;13(1):2332667. doi:10.1080/22221751.2024.2332667
- Puzelli S, Rossini G, Facchini M, et al. Human infection with highly pathogenic A(H7N7) avian influenza virus, Italy, 2013. Emerg Infect Dis. 2014;20(10):1745-1749. doi:10.3201/eid2010.140512
- Restori KH, Septer KM, Field CJ, et al. Risk assessment of a highly pathogenic H5N1 influenza virus from mink. Nat Commun. 2024;15(1):4112. Published 2024 May 15. doi:10.1038/s41467-024-48475-y
- Suttie A, Deng YM, Greenhill AR, Dussart P, Horwood PF, Karlsson EA. Inventory of molecular markers affecting biological characteristics of avian influenza A viruses. Virus Genes. 2019;55(6):739-768. doi:10.1007/s11262-019-01700-z
- The Texas Department of State health Services (DSHS). Health Alert: First Case of Novel Influenza A (H5N1) in Texas, March 2024. released 1 April 2024.
- U.S. Department of Agriculture. Federal and State Veterinary, Public Health Agencies Share Update on HPAI Detection in Kansas, Texas Dairy Herds. released 25 March 2024. 2024a.
- U.S. Department of Agriculture. USDA Confirms Highly Pathogenic Avian Influenza in Dairy Herd in Idaho. released 2 April 2024. 2024b.
- U.S. Department of Agriculture. USDA, HPAI Confirmed Cases in Livestock. Updated 3 July. 2024. 2024c.
- U.S. Department of Agriculture. USDA, Detections of Highly Pathogenic Avian Influenza (HPAI) in Livestock. Updated 20 December. 2024. 2024d.
- U.S. Department of Agriculture. USDA, Federal and State Veterinary Agencies Share Update on HPAI Detections in Oregon Backyard Farm, Including First H5N1 Detections in Swine. released 10 October. 2024. 2024e.
- U.S. Department of Agriculture. USDA, USDA Animal and Plant Health Inspection Service Shares Update on H5N1 Detection in Oregon Swine, Bovine Vaccine Candidate Progression. Released 6 November. 2024. 2024f
- U.S. Department of Agriculture. USDA, HPAI Confirmed Cases in Livestock. Updated 16 January. 2025. 2025a.
- U.S. Department of Agriculture. USDA, The Occurrence of Another Highly Pathogenic Avian Influenza (HPAI) Spillover from Wild Birds into Dairy Cattle. 2025b.
- U.S. Department of Agriculture. USDA, APHIS Confirms D1.1 Genotype in Dairy Cattle in Nevada. Updated 5 February. 2025. 2025c.
- UKHSA. UKHSA confirms rare case of bird flu A(H5N1) in the West Midlands region. 2025.
- WHO. 2019. Influenza at the human-animal interface Summary and assessment, 12 February to 9 April 2019.
- WHO. 2021. WHO information for the molecular detection of influenza viruses. Accessed.
- WHO. 2022. Assessment of risk associated with recent influenza A(H5N1) clade 2.3.4.4b viruses.
- WHO. Disease Outbreak News(DONs). As of 11 April 2023.
- WHO. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003-2024, 27 September 2024. 2024a.
- WHO. Disease Outbreak News(DONs) Avian Influenza A (H5N1) – Australia. released 7 June 2024. 2024b
- WHO. Influenza at the human-animal interface summary and assessment, 27 September 2024. 2024c.
- WHO. Disease Outbreak News (DONs). 2024d.
- WHO. Updated joint FAO/WHO/WOAH assessment of recent influenza A(H5N1) virus events in animals and people. 2024. 2024e.
- WHO. Updated joint FAO/WHO/WOAH public health assessment of recent influenza A(H5) virus events in animals and people. 2024f
- WHO, Influenza at the human-animal interface summary and assessment, 20 January 2025. 2025a
- WHO. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003-2025, 20 January 2025. 2025b
- WHO. Summary of status of development and availability of A(H5N1) candidate vaccine viruses and potency testing reagents. 28 February 2025. 2025c
- WHO. Summary of status of development and availability of A(H5) non–A(H5N1) candidate vaccine viruses and potency testing reagents. 28 February 2025. 2025d
- Rosenke K, Giffin A, Kaiser F, et al. Pathogenesis of bovine H5N1 clade 2.3.4.4b infection in Macaques. Nature. Published online January 15, 2025. doi:10.1038/s41586-025-08609-8
- WOAH. High Pathogenicity Avian Influenza (HPAI) –Situation report period covered: 17 February to 9 March 2023.
- WPRO. Avian Influenza Weekly Update Number 974. 22 November 2024.
- Yamada S, Suzuki Y, Suzuki T, et al. Haemagglutinin mutations responsible for the binding of H5N1 influenza A viruses to human-type receptors. Nature. 2006;444(7117):378-382. doi:10.1038/nature05264
- 磯田典和, 日尾野隆大, 迫田義博. 北海道における野鳥および野生哺乳動物からのH5N1亜型高病原性鳥インフルエンザウイルスの検出事例について. IASR Vol. 43 p259-260: 2022年11月号.
- 環境省. 2024. 動物園等における飼養鳥に関する高病原性鳥インフルエンザへの対応指針. 2024年11月14日
- 厚生労働省. 1998. 感染症の予防及び感染症の患者に対する医療に関する法律(平成10年法律第114号).
- 厚生労働省. 2006. 国内の鳥類における鳥インフルエンザ(H5N1)発生時の調査等について(平成18年12月27日付健感発第1227003号平成20年5月12日一部改正).
- 厚生労働省. 2008. 鳥インフルエンザ(H5N1)に係る積極的疫学調査の実施等について(平成18年11月22日付健感発第1122001号. 平成20年5月12日一部改正厚生労働省健康局結核感染症課長通知).
- 厚生労働省. 2024. 鳥インフルエンザ(H5N1)に関する積極的疫学調査の実施等について(依頼)(感感発1212第1号).
- 国立感染症研究所. 2015. 米国疾病予防管理センター(CDC)の提唱するインフルエンザウイルスのパンデミックリスクアセスメント.
- 農林水産省. 2023年~2024年シーズンにおける高病原性鳥インフルエンザの発生に係る疫学調査報告書. 2024a
- 農林水産省. 令和5年度 鳥インフルエンザに関する情報について. 2024b
- 農林水産省. 米国の乳牛における高病原性鳥インフルエンザウイルス(H5N1亜型)への感染事例について2024年4月3日. 2024c
- 農林水産省. 高病原性鳥インフルエンザ及び低病原性鳥インフルエンザに関する特定家畜伝染病防疫指針. 2020年7月1日(一部変更:2024年10月31日). 2024d
- 農林水産省. 2025. 令和6年度 鳥インフルエンザに関する情報について.
- 農林水産省. 家きん疾病小委員会及び高病原性鳥インフルエンザ疫学調査チーム検討会合同会合の提言を踏まえた防疫対策の徹底について. 2023年12月20日.
更新履歴
- 2023年12月8日
「海外渡航者が感染するリスク」内に体調不良による受信時の注意事項の啓発について追記 - 2024年4月17日
疫学的所見、ウイルス学的所見、リスクアセスメントについて更新
作成
感染症危機管理研究センター 村上裕子、太田雅之、竹前喜洋、嶋田聡、内木場紗奈、小林望、山本朋範、加藤美生、吉見逸郎、東良俊孝、影山努、関なおみ、齋藤智也
実地疫学研究センター 加藤博史、藤井英里、西野綾乃、島田智恵、砂川富正
FETP 25期 小野貴志、中村夏子
26期 小川順子、田才愛子、広瀬卓哉、中満智史
インフルエンザ・呼吸器系ウイルスセンター 渡邉真治、浅沼秀樹、長谷川秀樹
